Warum Bettwanzen-Spürhunde?
Unsere Spürhunde ermöglichen eine effektive Bettwanzenkontrolle. Wie machen sie das? Welche Voraussetzungen bringen Hunde für die Spürhundearbeit mit?
Während wir Menschen über ca. 20 Millionen Riechzellen verfügen, triumphiert der Hund (Border Collie, Labrador, DSH) mit ca. 220 Millionen. Die Riechleistung von Hunden ist derart sensitiv, dass sie selbst flüchtige organische Substanzen in niedrigster Konzentration von 1,14 ppt identifizieren können. Nehmen wir z.B. einen Teelöffel Zucker und lösen ihn in zwei Olympia-Schwimmbecken auf, so sind Hunde in der Lage, den Zucker herauszuriechen.
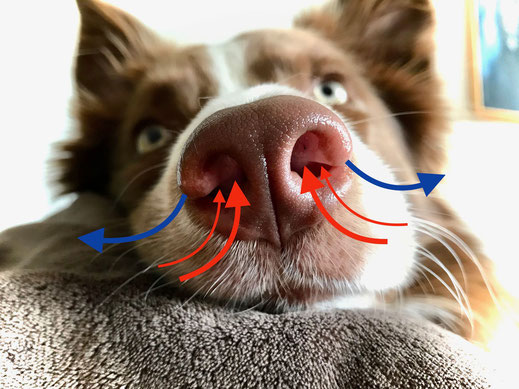
Spürhunde sind auf ein ganz bestimmtes Geruchsbild (= scent picture) trainiert, nach welchem sie im Einsatz gezielt suchen.
Hierfür setzen sie das aktive Schnüffeln in Form eines hochfrequentierten Ein- und Ausatmens ein. Je nach Schnüffelrate atmen Hunde bis zu 300 Mal pro Minute – 8-20 Mal ein, 1 Mal aus. Die Atemtechnik selbst ist äußerst effektiv: Eingeatmet wird durch die zentralen Nasenlöcher, ausgeatmet durch die seitlichen Schlitze. Beim Schnüffeln weicht die ausgeatmete Luft seitlich nach hinten und unten, wodurch erstens die Geruchsquelle vor der Nase nicht verwirbelt wird und zweitens in Bodennähe ein Unterdruck direkt vor der Nase entsteht. Hierdurch strömt die Umgebungsluft in Richtung Nasenlöcher und wird beim erneuten Einatmen der Nase zugeführt.
In der Nase selbst sorgen die Turbulenzen in den Nasengängen für eine erhöhte Riechleistung.
Darüber hinaus riechen Hunde in Stereo, d.h., den Nasenlöchern werden unabhängig unterschiedliche Luftproben zugeführt. Hunde haben sogar für jedes Nasenloch Präferenzen: Das rechte Nasenloch wird eher für das Erschnüffeln neuer oder stress- und erregungsauslösender Düfte benutzt, das linke Nasenloch dagegen bei neutralen, bekannten oder wenig aufregend eingestuften Gerüchen. Studien haben gezeigt: Nach wiederholtem Schnüffeln eines zuvor unbekannten Stoffs wechseln Hunde das Nasenloch.
Spürhunde identifizieren demnach den Zielgeruch mit dem linken Nasenloch!
Die Nervenbahnen fürs Riechen kreuzen sich übrigens nicht! Informationen via rechtes Nasenloch gehen in die rechte Gehirnhälfte, Informationen via linkes Nasenloch in die linke Gehirnhälfte.
Die enorme Riechleistung der Hunde beruht zudem auf dem Jacobsonschen Organ (Vomeronasalen Organ), mit dem Duft- und Botenstoffe identifiziert werden – all dies erlaubt eine schnelle und zuverlässige Detektion.
- Concha, A. et al. (2014). Using sniffing behavior to differentiate true negative from false negative responses in trained scent-detection dogs, Chem Senses, 39(9), 749–754.
- Horowitz, A. (2009). Inside of a dog: What dogs see, smell, and know. New York: Scribner.
- Jenkin, E.K. et al. (2018). When the nose doesn’t know: Canine olfactory function associated with health, management, and potential links to microbiota. Front Vet Sci, 5(56), 5–56.
- Settles, G. S. (2005). Sniffers: Fluid-Dynamic Sampling for Olfactory Trace Detection in Nature and Homeland Security. Journal of Fluids Engineering, 127(2), 189–218.
- Wolf, A. (2016). Untersuchung des Einflusses der Alterung menschlicher Geruchsspuren auf die Ausarbeitung der Fährten durch Personensuchhunde. (Dissertation, Tierärztliche Hochschule Hannover, 2016). Gießen: DVG Service GmbH.
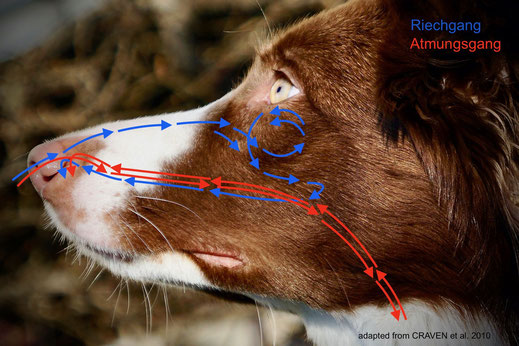
Bei Hunden ist die nasale Strömungsdynamik (Fluiddynamik) anatomisch durch den sowohl kompakten als auch komplexen, multifunktionalen Aufbau der Nasenhöhle bedingt. Computergestütze Berechnungen des inspirativen Atemflusses während des Riechens/Schnüffelns zeigen, dass Geruchs- und Atmungsluftströme im Wesentlichen zwei getrennte Phänomene mit ganz eigenen Strömungsbahnen sind.
Die für das Riechen ausschlaggebende obere Bahn (Riechgang, in der obigen Abb. in Blau) ermöglicht den direkten Transport von Luft zur olfaktorischen Region, und zwar unabhängig von der Atmung. Nachdem die eingeatmete Luft im Nasenvorhof durch Turbulenzen vermischt wurde, fließt ein geringer Teil der Luft mit hoher Geschwindigkeit in einer schmalen Passage, dem dorsalen Nasengang (meatus nasi dorsalis), durch das Dach zum hinteren Bereich der Nasenhöhle, zur olfaktorischen Region im Nasengrund. In diesem hintersten Bereich der Nasenhöhle wendet sich die Luft um 180º und flutet langsam das mit Riechschleimhaut und Nervenzellen ausgekleidete, stark gefaltete Siebbeinlabyrinth. Dieses Riechfeld umfasst eine Größe von ca. 170 cm2, während wir Menschen uns mit mickrigen 10 cm2 begnügen müssen. Eine knöcherne horizontale Platte (lamina transvera) trennt diese Riechregion vom Nasenrachen und bildet einen olfaktorischen Recessus, eine Art Nische, die dafür sorgt, dass die Luft nicht nur länger, sondern sogar während des Ausatmens verweilen kann. Auf diese Weise werden mehr Geruchsmoleküle von der Riechschleimhaut gebunden und damit gelangen mehr Geruchspartikel in Kontakt mit den Riechrezeptoren, womit die Identifizierung der Geruchsstoffe optimiert wird. Der Hund riecht also selbst während des Ausatmens!
Danach fließt die Luft zum Nasenrachen, von wo aus sie die Nasenhöhle über die untere Bahn (Atmungsgang, in der obigen Abb. in Rot), den ventralen Nasengang (meatus nasi ventralis) am Nasenboden entlang, durch die Nasenlöcher wieder verlässt.
Beim normalen Atmen mit einem entspannten, nicht ausgedehnten Nasenloch sorgt das Aufspalten in Geruchs- und Atmungsluftstrom dafür, dass etwa 12-15 % der inspirierten Luft die olfaktorische Region erreicht. Beim Schnüffeln allerdings mit einer erhöhten physiologischen Nasenlochbewegung liegt der Prozentsatz höher. Mit dem erhöhten Volumen der eingeatmeten Luft nimmt natürlich auch die Menge der Geruchsmoleküle zu, die die Riechregion erreicht, womit die Riechleistung gesteigert wird.
Auch das vomeronasale Organ (Jacobsonsche Organ) im Nasenboden und zugleich am Gaumendach des Hundes zählt zum olfaktorischen System und wird übrigens ebenso nicht von der normalen Atmung beeinflusst. Mit ihm werden Pheromone wahrgenommen, aber mittlerweile ergeben sich Hinweise, dass auch die olfaktorische Region am Ende des Riechgangs für die Pheromondetektierung zuständig ist.
- Craven, B. A., Paterson, E. G. and Settles, G. S. (2010). The fluid dynamics of canine olfaction: unique nasal airflow patterns as an explanation of macrosmia. J. Roy. Soc. Interface 7, 933-943.
- M. J. Lawson, M. J., Craven, B. A., Paterson, E. G. and Settles, G. S. (2012). A computational study of odorant transport and deposition in the canine nasal cavity: implications for olfaction. Chem. Senses 37, 553–566.
- Thomas P. Eiting, Timothy D. Smith, J. Blair Perot, Elizabeth R. Dumont (2014). The role of the olfactory recess in olfactory airflow. J. Exp. Biol. 217, 1799-180.
- Wolf, A. (2016). Untersuchung des Einflusses der Alterung menschlicher Geruchsspuren auf die Ausarbeitung der Fährten durch Personensuchhunde. (Dissertation, Tierärztliche Hochschule Hannover, 2016). Gießen: DVG Service GmbH.